
- The twentieth century was indeed the century of Modern Medicine with tremendous strides made in the understanding and control of infectious diseases, as well as the introduction of life-saving antibiotics and vaccines. Unfortunately, along with these advances came the perils of genetic engineering, the increasing threat of newly emerging viruses, biowarfare, and bioterrorism
- Despite these scientific achievements, the cause of cancer remains a mystery. Scientists suspect genetic susceptibility, possible cancer-causing viruses, and environmental factors might play a role in some cancers, but none of these factors explain why millions of people die yearly from a variety of malignancies.

William Russell 1852-1940, as pictured in
The British Medical Journal, August 24, 1940.- How could scientists put men on the moon, but remain so ignorant about cancer and its origin? How can the infectious causes of tuberculosis, leprosy, syphilis, smallpox, polio, malaria, and other viral and bacterial and parasitic diseases be understood, but the cause of cancer be unknown? Could the cause of cancer conceivably be an infectious agent that has been overlooked, ignored, or unrecognized by medical doctors in the twentieth century? Could the germ of cancer be hidden in the Russell body? - a large microscopic form known to every pathologist for over a century!
- William Russell (1852-1940) and "the parasite of cancer"
- On December 3, 1890 William Russell, a pathologist in the School of Medicine at the Royal Infirmary in Edinburgh, gave an address to the Pathological Society of London in which he outlined his histopathologic findings of "a characteristic organism of cancer" that he observed microscopically in fuchsine-stained tissue sections from all forms of cancer that he examined, as well as in certain cases of tuberculosis, syphilis and skin infection.
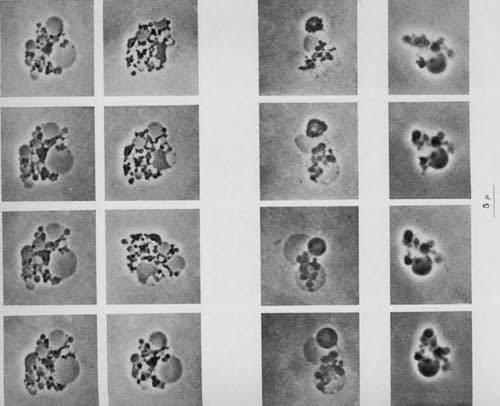
 The parasite was seen within the tissue cells (intracellular) and outside the cells (extracellular). The size of Russell's parasite ranged from barely visible, up to "half again as large as a red blood corpuscle." The largest round forms were easily seen microscopically. The large size of some of these bodies suggested a fungal or yeast-like parasite. Russell provisionally classified the parasite as a possible "blastomycete" (a type of fungus); and called the forms "fuchsine bodies" because of their bluish-red staining qualities.
The parasite was seen within the tissue cells (intracellular) and outside the cells (extracellular). The size of Russell's parasite ranged from barely visible, up to "half again as large as a red blood corpuscle." The largest round forms were easily seen microscopically. The large size of some of these bodies suggested a fungal or yeast-like parasite. Russell provisionally classified the parasite as a possible "blastomycete" (a type of fungus); and called the forms "fuchsine bodies" because of their bluish-red staining qualities. - Microbiology was still in its infancy in Russell's era, and it was generally thought that each microbe could only give rise to a single disease. Thus, the idea of a cancer germ (especially one that could also be identified in TB and syphilis) was received cautiously. Nine years later in 1899, in yet another report on "The parasite of cancer" appearing in The Lancet (April 29), Russell admitted that finding cancer parasites in diseases other than cancer was indeed a "stumbling block." By this time a considerable number of scientists concluded that Russell bodies were merely the result of cellular degeneration of one kind or another. Furthermore, no consistent microbe was cultured from tumors; and the inoculation of these microbes into animals produced conflicting and often negative results.
- Russell was trained as a pathologist, not as a microbiologist, and he avoided getting into the bacteriologic controversies regarding various microbes grown from cancer. He simply concluded, "It seems almost needless to add that there remains abundant work to be done in this important and attractive field."
- After three years' work at the New York State Pathological Laboratory of the University of Buffalo, Harvey Gaylord confirmed Russell's research in a 36 page report titled "The protozoon of cancer", published in May, 1901, in the American Journal of the Medical Sciences. Gaylord found the small forms and the large sacs characteristic of Russell bodies in every cancer he examined. Some large spherical bodies were four times the diameter of a leukocyte (white blood cell). Red blood cells measure about 7 micron in diameter and leukocytes are 2 to 3 times larger than red blood cells. Thus, some of the bodies that Gaylord observed attained the amazing size of around 50 micron in diameter. In addition, he found evidence of internal segmentation within the larger bodies "after the manner recognized in malarial parasites." The tiniest forms appeared the size of ordinary staphylococci.

Russell bodies in a lymph node of Hodgkin's disease.
Gram's stain, magnified 1000 times, (in oil).- Russell's 1899 paper ended his writings of a cancer parasite, but his discovery quickly became known to pathologists as Russell bodies. These bodies continue to fascinate researchers and physicians (like myself) up to the present time.

Solitary "giant" Russell body in a lymph node of Hodgkin's disease
(cancer), magnified 1000 times. Gram's stain, magnified 1000 times.- When Russell died at the age of 89 in 1940, the British Medical Journal published a large obituary noting that he was universally respected and embued with the dignity and highest ideals of his profession, and that he had served at one time as President of the Royal College of Physicians. No mention was made of his "parasites" or his "bodies", except to remark that "in his earlier years Russell devoted much time to the study of the cancer cell." Similarly, a large obituary appeared in the Edinburgh Medical Journal along with a full-page photo. His published books on Clinical Methods and widely read texts on circulation and gastro-intestinal diseases were cited, but not a word about his discovery in cancer.
- The heresy of "the cancer microbe"
- By the early part of the twentieth century the top cancer experts had all rejected so-called "cancer parasites" as the cause of cancer. The most influential physician to speak against it was James Ewing, an American pathologist and author of the widely-read textbook, Neoplastic Diseases. In 1919 Ewing wrote that "few competent observers consider it (the parasitic theory) as a possible explanation in cancer." According to Ewing and other authorities, cancer did not act like an infection. Therefore, microbes could not possibly cause cancer. He concluded, "The general facts of the genesis of tumors are strongly against the possibility of a parasitic origin."
- As a result, the parasitic theory was totally discarded and few doctors dared to contradict Ewing's dogma by continuing to search for an infectious agent in cancer. Nevertheless, a few die-hard physicians remained convinced microbes were at the root cause of cancer and wrote about it convincingly in medical journals. The long history of this research is recorded in my book, The Cancer Microbe (1990) and anyone with internet access can do a Google search (type in "cancer microbe") and obtain a wealth of information on the microbiology of cancer. Another excellent history of cancer microbiology and the suppression of this controversial research is contained in David Hess' Can Bacteria Cause Cancer? (1997).

Photo #5: Larger Russell bodies (RB) in a lymph node showing non-cancerous "reactive lymph node hyperplasia" from a fatal case of AIDS. The arrow points to nearby bacterial-sized intracellular coccoid smaller forms from which the Russell bodies are derived. Fite (acid-fast) stain, magnified 1000 times.- In the 1920s James Young, an obstetrician from Scotland, repeatedly grew pleomorphic (having many forms) bacteria from various cancers. The microbes had a "specific life cycle" and "spore stages" comprised of exceedingly tiny and barely visible spores. In the laboratory these tiny spores transformed into larger coccoid (round) forms, rod-forms and yeast-like forms (similar in size to Russell bodies). John Nuzum, a Chicago physician, reported a pleomorphic coccus he repeatedly isolated from breast cancer. The tiniest forms were virus-like and passed through a filter designed to hold back bacteria.
- In 1925 Northwest Medicine published two papers by Michael Scott, a Montana surgeon who learned about the cancer microbe in TJ Glover's lab in 1921. Scott's microbe was similar to Young's. The parasite had a life cycle composed of three stages: a coccus, a rod, and a "spore sac" stage. Scott believed cancer was an infection like tuberculosis and attempted a vaccine treatment, but his treatment methods were quickly suppressed by the medical establishment.
- In the 1930s in Germany the controversial Wilhelm von Brehmer described microbes in the blood of cancer patients, evoking the wrath of his scientific colleagues and prompting an intervention by Adolf Hitler. (See Proctor's The Nazi War on Cancer [1999]) Georges Mazet, a French physician, also found pleomorphic bacteria in Hodgkin's disease in 1941. Hodgkin's is a type of lymphoma cancer involving the lymphatic system. Mazet later reported similar acid-fast (red staining) bacteria in many different kinds of cancer, including leukemia.
- In the 1950s, 60s, and 70s, a quartet of women further refined the microbiology of cancer, emphasizing the extreme pleomorphism of the organism and its detection in tissue with the acid-fast stain. The published research of Virginia Livingston, Eleanor Alexander-Jackson, Irene Diller and Florence Seibert, is essential reading for the most updated understanding of the microbiology of cancer.
- In the late 1970s Guido Tedeschi and other Italian microbiologists at the University of Camerino discovered "granules" in the red blood cells of healthy and ill people that turned out to be bacteria that could be cultured in the laboratory. Some of the staphylococcal and corynebacteria-like bacteria cultured from the red blood cells were acid-fast and cell wall-deficient, a staining and growth characteristic shared with the cancer microbe. This research has been confirmed by newer studies suggesting that bacteria reside in blood from healthy as well as sick individuals. These findings of tiny blood bacteria (nanobacteria) provide further evidence to support the theory that microbes can cause cancer.
- Some other well-known scientists in the field of cancer microbiology include Gunther Enderlein, Royal Raymond Rife, Gaston Naessens and Wilhelm Reich. All have web sites devoted to their cancer research.
- Russell bodies and their Origin
- More than a century has passed since Russell's discovery and although electron microscopes (which have been used since the 1950s) have the ability to magnify objects tens of thousands of times, the significance and function of his bodies still remains unknown.
- What is well-known is that Russell bodies can be found, not only in cancer, but in the majority of inflamed tissues throughout the body. Distinguishing large Russell bodies from actual fungal forms of Blastomyces can still be difficult, particularly when a pathologist encounters a true case of fungal infection due to Blastomyces.
- In 1954 RG White, in "Observations on the formation and nature of Russell bodies", produced Russell bodies in animals by injecting them with different species of bacteria. He then studied the ensuing development of these bodies in the spleen, lymph nodes and plasma cells of the injected animals. Plasma cells are specialized forms of white blood cells that normally produce antibodies.
- EM Schleicher, in his 1965 paper on "Giant Russell bodies", discusses the various theories of origin. Possibilities include origin from the lymphocyte, origin in plasma cells with later degeneration, origin from the mitochondria of cells, and even an origin from a red blood cell (erythrocyte) swallowed up by a plasma cell.
- Most researchers currently believe Russell bodies are essentially immunoglobulins (proteins that acts as antibodies), but an electron microscopic study by SM Hsu et al. in 1981 has cast some doubt on this belief.
- None of these studies mention the possibility that Russell bodies might represent unusual large growth forms of bacteria. However, if Russell bodies prove to be tiny intracellular microbes that grow and enlarge within leukocytes, it would be natural to expect these white blood cells (especially the plasma cell) to produce an antibody attack against these invading organisms, resulting in the production of immunoglobulin-coated cells and organisms.
- Bacterial transformation into Giant forms (L-form "large bodies')
- There are many different kinds of bacteria but only one type that has been consistently observed and studied in cancer for over a century. The cancer microbe has many forms, some of which appear as ordinary staphylococci or larger yeast-like forms that further enlarge to the size of Russell bodies. As mentioned, some Russell bodies enlarge to truly gigantic proportions, one hundred times the diameter of small cocci. One can liken this growth potential to an empty balloon that is then blown up to full-size. In addition, the microbe has exceedingly small filterable submicroscopic forms approaching the size of viruses, visible only by use of the electron microscope.
- Scientists who have extensively studied the cancer microbe claim it most closely resembles the type bacteria that cause tuberculosis and leprosy- the so-called mycobacteria. Mycobacteria are closely related to fungi; and some microbiologists claim mycobacteria are essentially derived from the "higher" fungi. "Myco" in Greek means fungus. Ergo, mycobacteria are considered fungus-like bacteria.

Photo #6: Extremely large "super-giant-sized" solitary Russell body in the skin of "cutaneous lupus erythematosus", a so-called "collagen disease." The perfectly round shape, except for one area, suggests this large body is developing inside a cell that is readly to burst. Kinyoun's (acid-fast) stain, magnification x 1000.- During the 1960s microbiologist Louis Dienes popularized the terms "cell wall-deficient" and "L form" to encompass bacterial growth stages that exist at one extreme as small filterable virus-sized forms, and at the opposite extreme as large (50 micron or larger) spherical forms that he termed "large bodies." These so-called large bodies are what I believe Russell bodies represent.
- It must be understood that microbes are partially "classified" in microbiology according to size. Viruses are submicroscopic and cannot be visualized with an ordinary light microscope. Unlike bacteria, viruses can only replicate inside a cell. Bacteria can be seen microscopically, but smaller submicroscopic and filterable bacterial forms (now known as nanobacteria) are also known. Fungi and yeast forms are much larger than bacteria, and "mold" can obviously be seen with the naked eye.
- Larger Russell bodies are indeed similar in size to certain spore forms of fungi. However, what is generally not appreciated is that bacteria can grow into fungal-sized large bodies, depending on certain laboratory conditions. Thus, bacteria in this form can easily be mistaken for fungi and yeast organisms.
- Giant-sized L-forms greatly resemble large-sized Russell bodies. The century-old history of research into atypical growth forms of bacteria is reviewed in Lida Mattman's seminal text, Cell Wall Deficient Forms: Stealth Pathogens (1993). A knowledge of this somewhat esoteric branch of microbiology is essential to understand the proposed microbiology of cancer.
- The most impressive electron microscopic photographs I have ever observed of cell wall-deficient L-forms of mycobacteria were taken by the late C Xalabarder of Barcelona. In a series of papers and books (1953-1976) published in Spanish (with English-language summaries) by the Publicaciones del Instituto Antituberculoso "Francisco Moragas", Xalabarder totally transformed my concept about how tuberculosis-causing mycobacteria reproduce and grow and drastically change their appearance. In medical school we were taught that "simple" bacteria simply divide in two equal halves by "binary fission". However, nothing could be further from the truth, and it is only by a refutation of this simplistic concept that a serious study of the microbiology of cancer can be undertaken.
- Tuberculosis and Cancer
- Because cancer is produced by a microbe similar to the bacteria that cause TB, much can be learned from experiments like those performed by Xalabarder in 1967. Using "atypical mycobacteria" grown from TB patients who had taken long courses of drug therapy, Xalabarder then injected these bacteria into guinea-pigs and rabbits. Amazingly, he was able to experimentally produce lesions which microscopically resembled cancer! He also produced experimental lesions characteristic of so called "collagen disease"- a type of lesion seemingly unrelated to cancer.
- During the 1960s I discovered unusual pleomorphic acid-fast bacteria in a collagen disease called scleroderma, and later in another collagen disease called lupus erythematosus. The germs I grew from these patients closely resembled scleroderma microbes that were reported by Virginia Livingston in 1947, and which subsequently led to her discovery of similar acid-fast microbes in cancer.
- In 1969 Xalabarder manipulated different developmental stages of TB bacteria and inoculated them into one thousand guinea pigs. In the process, he produced the microscopic picture of sarcoidosis in the animals. Sarcoidosis is a human disease closely related to TB but one in which TB germs cannot be found. Xalabarder's most impressive sarcoid lesions were produced by inoculating sputum specimens from TB patients who "converted", meaning that their TB bacteria could no longer be cultured from their sputum. Controversy over the cause of sarcoidosis is still not settled, although I reported bacteria similar to cancer microbes in this disease in the 1980s.
- The most spectacular electron microphotographs of cell wall-deficient mycobacteria are presented in Xalabarder's L-forms of mycobacteria and chronic nephritis (1970). In the earliest growth stages of mycobacteria in culture the smallest elements appear as tiny submicroscopic forms visualized only with the electron microscope. These filterable forms of tuberculosis bacteria - the so-called "tuberculosis virus"- have been known to cause cancer in animals since the 1920s. By adding antibiotics to the lab culture media Xalabarder was able to induce many unusual growth forms of tuberculosis bacteria. Using serial images, he was able to trace the development of these tiny submicroscopic forms up to the size of ordinary cocci - and then up to the size of "large body" forms reaching and even surpassing the size of red blood cells. Some of the large bodies of mycobacteria also exhibit internal structure, similar to what Gaylord noted in his Russell body research.
- Cancer and Bacteria
- Although the idea of a cancer microbe is medical heresy, there is ample data to show that cancer patients are highly prone to bacterial infection. A PubMed computer search (http://www.ncbi.nlm.nih.gov/entrez/query.fcgi)
- of "bacteria + cancer" elicits 49, 244 citations contained within 2,463 web pages. According to a 2003 article by Vento and Cainelli, patients with cancer who are undergoing chemotherapy are highly susceptible to almost any type of bacterial or fungal infection.
- Why are physicians, and especially pathologists and bacteriologists, so unaware, so disinterested, or so antagonistic to credible cancer microbe research? Why have pathologists failed to consider Russell bodies as large forms of bacteria?
- For over 30 years I studied various forms of cancer and skin diseases "of unknown origin", as well as autopsy cases of cancer, lupus, scleroderma, and AIDS. In all these diseases I was able to detect bacteria, although pathologists would never mention bacteria in any of their official biopsy reports. In my experience, they simply could not conceive of cancer and collagen disease (and AIDS) as a bacterial infection, nor did they seem to be aware of bacteriology reports pertaining to "large bodies" and pathologic effects produced by the "tuberculosis virus." In short, they were trained to see and report only the typical rod-shaped acid-fast (red-stained) "typical" form of mycobacteria, , but they were not trained to look for or to recognize other growth forms of the same bacteria that might be hidden in their pathologic tissue specimens.
- Photo #7: Pleomorphic growth forms (L-forms) of tuberculosis mycobacteria photographed with an electron microscope. Note the darker staining tiny coccal forms (similar in size to ordinary staphylococci) and the larger clear balloon-sized "ghost" forms similar in size and shape to Russell bodies found in tissue. These forms are all characteristic of "cell wall-deficient bacteria" and totally unlike the well-known "typical" acid-fast rod forms of Mycobacterium tuberculosis. Reproduced from L-forms of Mycobacteria and Chronic Nephritis (1970), by Dr. C. Xalabarder P., page 51.
- When objects like Russell bodies are observed in a wide variety of diseases and in "normal" tissue, the significance is lessened. Doctors expect "normal" tissue to be free of microbes. I suppose they also conclude that Russell bodies cannot be an infectious agent because it would be impossible for an infectious agent to appear in so many different kinds of diseases and in so many different forms of cancer.
- For most of the last century stomach ulcers were thought to be non-infectious because pathologists could not identify bacteria in the ulcers and because doctors believed bacteria could not live in the acid environment of the stomach. This thinking all changed gradually after 1982 when Barry Marshall, an Australian physician, proved most stomach ulcers were caused by a microbe called Helicobacter pylori, which could be identified microscopically with special tissue staining techniques in ulcer tissue. On the other hand, many people normally carry this stomach microbe without any ill effects. Not surprisingly, pathologists are now reporting numerous Russell bodies in plasma cells in some ulcer patients, giving rise to a previously unrecognized tissue reaction called "Russell cell gastritis."
- Russell bodies and bacteria
- When bacteria are threatened by the immune system or by antibiotics they may lose their cell-wall and assume a different growth form that renders them less susceptible to attack by the immune system. Some Russell bodies elicit little or no inflammatory cell response. This lack of cellular response is yet another reason why physicians have a hard time believing Russell bodies could be microbes.
- I have observed the largest and most complex Russell bodies in tissue where there was almost a total lack of inflammation. My photographs of such "large bodies", some with obvious internal structure, that I observed in patients with scleroderma and pseudoscleroderma, were published in the American Journal of Dermatopathology in 1980. The first case of fatal scleroderma I studied in 1963 had numerous "large bodies" in the fat layer of the diseased skin that were unlike anything ever seen in dermatology. The patient had been hospitalized for pulmonary tuberculosis 7 years before developing scleroderma. The mystery of these "yeast-like" bodies deep in his skin was solved years later when I first learned about the existence of "large body" forms of Mycobacterium tuberculosis. When this patient died, Mycobacterium fortuitum, an "atypical" form of mycobacteria was cultured from his scleroderma tissue.
- Bacteria are vital for our survival. They are hardy and the bacteria we carry will surely outlive us. The bacteria that cause cancer are the "simple" bacteria we carry with us. The cancer microbe is not an exotic microbe nor a rare one. However, bacteria can change form as the environment in our bodies changes. There is indeed a delicate balance between our bacteria and our immune system which allows these bacteria to live in harmony with us.
- But when dis-ease occurs these microbes become aggressive, giving rise to a host of diseases, some of which are cancerous, and others that are inflammatory, degenerative, or simply transitory. Another reason for physicians to doubt that a single type of germ could cause such a variety of pathologic effects.
- Bacteria are ubiquitous and so are Russell bodies. And if Russell bodies prove to be bacteria, the reason for this becomes obvious.
- The Russell body and the origin of cancer
- In 1981 King and Eisenberg's article on "Russell's fuchsin body: 'The characteristic organism of cancer' " appeared in the American Journal of Dermatopathology. They reconfirmed that "Russell bodies have now been shown to be immunoglobulins." They remarked that Russell was not the first to describe them; and that similar bodies were reported by Cornil and Alvarez in rhinoscleroma five years earlier in a French journal in 1885. Declaring it ironic that these "bodies should bear the name of a man who so thoroughly misunderstood them", the authors ended by stating: "Hence, when the term Russell body is used today, one should be aware that the eponym is as inaccurate as was Russell's perception of their significance."
- Unlike King and Eisenberg, I believe Russell was right on the mark. There is a parasite in cancer. It has been studied and reported by various scientists throughout the world for many decades, and a wealth of scientific information on the cancer microbe is available in medical libraries. For those with Internet capability, the words "cancer microbe" typed into Google.com will give instant access to a treasure trove of information on the subject.
- There is no secret to cancer. In my view, the cause is staring us right in the face in the form of the Russell body. William Russell understood very well in the nineteenth century what medical science in the twenty-first century has yet to discover.
As a physician-dermatologist I have studied various aspects of the cancer microbe for over 30 years. In my book, The Cancer Microbe (Aries Rising Press, 1990), I recount a century of research by various scientists who have documented the reality and importance of bacteria associated with cancer. Despite a wealth of information on the microbiology of cancer, this body of work has been largely ignored.
Why would medical science overlook the finding of bacterial elements in cancer, particularly when the treatment of advanced cancer is often abysmal and when the cause (or causes) of many types of cancer remain unknown? If and when the bacterial cause of cancer is widely accepted, it will be left to medical historians to determine why the medical community failed to recognise cancer bacteria. At the present time, it is fair to say that most physicians are either unaware of cancer microbe research, or ignore the published findings, or are openly hostile to this research.
Unfortunately, medical doctors are limited by dogma about cancer-associated bacteria that eliminated a bacterial cause for cancer a century ago. In the late nineteenth century, when the bacterial cause of many infectious diseases was discovered, it was decided that cancer did not act like an infectious or contagious disease, and therefore it was concluded that bacteria were not causative.
Although a few scientists later found highly unusual and pleomorphic bacteria, these bacteria were simply dismissed as “contaminants” – or as microbes that had “secondarily infected” cancerous growths. Furthermore, there was no single or consistent type of microbe found, and animals experimentally infected with cancer microbes did not give develop cancer. Thus, decades before the rise of virology and molecular biology, and at a time when “mycoplasma” forms of bacteria were not known, the medical establishment concluded that bacteria were not involved as a cause of cancer in any way. This conclusion has coloured medical thought about cancer to this day.
Historically, it took centuries for doctors to recognise microbes as the cause of any disease. By the use of lenses, germs were discovered 200 years before physicians finally understood that microbes were capable of causing disease. For two centuries the dogma was that those exceedingly tiny “animacules” could not possibly be a threat to a grown person.
Once something becomes dogma in medical science, it is very difficult to change medical thinking. Ordinarily, infectious bacteria can be easily recognised in disease because they can be seen microscopically in tissue sections from disease states. Sometimes careful “special staining” of tissue sections is necessary to make microbes more visible and more easily identifiable. (In cancerous tissue, the cancer microbe is most easily viewed with an “acid-fast” tissue stain, like the special stain employed to identify the mycobacteria that cause tuberculosis and leprosy).
In this so-called modern era of medical science, one would think it impossible for disease experts to overlook disease-causing bacteria. However, when a new and deadly lung disease broke out among legionnaires in Philadelphia in July 1976, two hundred twenty-two people became ill and thirty-four died. The cause of the lung disease remained a medical mystery for over five months. Bacterial infection was ruled out when all tests were reported as negative. Fortunately, one astute and careful microbiologist finally discovered bacteria. Joe McDade at the Leprosy Branch of the CDC, was able to detect “unusual bacteria” in guinea pigs experimentally infected with lung tissue from the dead legionnaires. Further modification of bacterial culture methods finally allowed the isolation of causative bacteria, now known as Legionella pneumophila.
Yet another modern example of dogma-defying research is provided by recent studies proving that bacteria (Helicobacter pylori) are a common cause of stomach ulcers, which can eventually lead to stomach cancer and lymphoma. When I went to medical school, stomach ulcers were thought to be due to stress, lifestyle, or improper diet, and it was not uncommon to send ulcer patients to psychiatrists for analysis.
For a century, physicians refused to believe that bacteria could cause ulcers because they thought bacteria could not live in the acid environment of the stomach. In 1982 a researcher, who was unable to convince his colleagues that bacteria could cause ulcers and gastritis, actually proved his case by drinking a culture of H. pylori. When he rapidly became ill with stomach symptoms, he admitted himself to the hospital where these bacteria were found to be associated with his gastric disease. It also turned out that these bacteria could indeed be detected in the stomach lining of stomach ulcers, but only when the tissue was stained in a special way to detect the bacteria. The CDC now claims that H. pylori causes more than 90% of duodenal ulcers and 80% of gastric ulcers. Approximately two-thirds of the world’s population is infected with these microbes.
The present experience with ulcer-causing microbes proves that bacteria can indeed pop up in diseases where they are least expected. Such a caveat is appropriate for doctors who think they know everything about cancer and who pooh-pooh all aspects of cancer microbe research.
One perennial complaint about the so-called cancer microbe is that is pleomorphic. For some reason, the idea that a proposed cancer germ could have more than one form is a threat to doctors and some microbiologists. Indeed the cancer germ has been described as having a virus like and fungus-like, as well as mycoplasma-like phase. Such a “life cycle” is deemed nonsense and microbiologic heresy.
The many guises of the pleomorphic cancer microbe was studied extensively in the 1960s and 70s by four remarkable women scientists: Virginia Livingston (a physician); Eleanor Alexander-Jackson (a microbiologist); Irene Diller (a cytologist); and Florence Seibert, a chemist, tuberculosis expert, and inventor of the tuberculin skin test. Their individual and collaborative studies are essential reading to understand the proposed microbiology of cancer.
This research clearly indicated that cancer microbes are best detected by special tissue testing (similar to those used in tuberculosis and leprosy research). And that the cancer germ has some similarity to pleomorphic tuberculosis germs.
In all its many forms the tuberculosis microbe is certainly pleomorphic. (See the work of mycoplasma expert Lida H. Mattman.) The bacteria that cause TB are known as “mycobacteria”. Some forms of the bacillus are round “coccoid” forms; other forms are more typically “acid-fast” and “rod” forms. All mycobacteria form a phylogenetic link or bridge between the bacteria and the “higher” fungi. “Myco” is Greek for fungus. Ergo, myco-bacteria.
Under appropriate conditions, bacteria can lose their cell wall and become amorphous, smaller, highly pleomorphic “cell-wall deficient forms.” Under suitable conditions, mycoplasma can enlarge to giant-sized forms (“Large bodies”) resembling fungal and spore-like forms. It is vital to be aware of and to recognise such unusual and hard-to-detect forms in tissue microscopic sections because, in my experience, this mycoplasmal form is the form the cancer microbe takes inside the body in human disease. Due to their small size, Mycobacteria form a bridge between (larger) bacteria and smaller) viruses. Microbiologists love to separate (and classify) viruses, bacteria, mycoplasma, and fungi, as distinct entities. In fact, there is interplay between all of them. It is well-known that bacteria can be infected with viruses. Nevertheless, scientists cannot seem to understand how microbes can change into virus-like, mycoplasma-like and fungus-like infectious agents.
Because the cancer microbe is related to the bacteria that cause tuberculosis, it is helpful to compare the microbiology of cancer with what we know about the microbiology of mycobacteria and their production of various forms of clinical TB.
Over the past half-century we have learned that TB is not always caused by the same identical germs. TB infections of the lung may be caused by various “atypical” mycobacteria that are not identical to the common Mycobacterium tuberculosis. Also some atypical mycobacteria have been discovered in various disease states that are not considered tuberculosis. Thus, there is no reason to expect all cancer-associated bacteria to be exactly the same germ.
Furthermore, just as everyone who harbours H. pylori does not develop stomach ulcers, we should not expect all “cancer microbes” to produce cancer. Also it is not unreasonable to consider that cancer microbes have the potential to produce disease states that are not considered cancer.
For many years I identified cancer microbes in a variety of disease states. In The Cancer Microbe, I show photomicrographs of cancer microbes in “autoimmune” diseases such as scleroderma, in AIDS-related Kaposi’s sarcoma, in enlarged lymph nodes in AIDS, in breast cancer, in lymphoma and Hodgkin’s disease, in a lung disease called interstitial pneumonitis, in sarcoidosis, in an immunoblastic sarcoma and even in a skin cancer.
Not everyone who becomes infected with TB germs develops clinical tuberculosis. People can harbour the TB germ without ever becoming ill. The same is true for cancer microbes. Not everyone who carries them develops cancer.
According to Virginia Livingston, the microbe is “ubliquitous.” It is found in various disease states and also can be found normally. This is a difficult for some medical doctors to believe because of the idea that an infectious agent must always infect. Livingston infuriated the scientific establishment by naming the cancer microbe “Progenitor cryptocides” – meaning “hidden killer”). She claimed the microbe was present in every cell. Due to its biochemical peculiarities, the organism was responsible for initiating life and for healing of tissue; and was the microbe ultimately responsible for eventual degeneration and death of all life. Such ideas, of course, are at odds with medical thought. However, my own studies have suggested that the cancer microbe is indeed ubiquitous and indestructible, which is further reason why it should be taken seriously, particularly in diseases that are poorly understood, like cancer and “diseases of unknown etiology.”
Most importantly, cancer microbes are significant because they can be identified in the cancerous tissue in various forms of cancer. A few of these microbes can be seen in “normal” tissue, but strikingly larger numbers can be seen in the areas of the tumour. These microbes can be identified in “pre-cancerous” conditions, suggesting that these germs are present before the actual induction of the cancer. Furthermore, when cancer is “cured” by radiation and chemotherapy, the microbe can still be found in the damaged, previously cancerous areas.
The reason we cannot “cure” cancer is that we cannot stop the destruction caused by these “hidden” and “unrecognised” bacterial elements. The reason antibiotics do not work well in cancer is because the microbes (in the mycoplasmal phase inside the body) are not susceptible to antibiotics.
In cancer research, there is controversy as to whether cancer is one disease or many. For instance, could breast cancer and lung cancer and prostate cancer all be caused by the same agent. This would be deemed highly improbable, but if cancer microbes were shown to be associated with all three forms of cancer, the possibility that all three kinds of cancer might be related becomes more possible.
When Livingston and colleagues injected cancer microbes into animals and chickens, some developed cancer, some developed degenerative and proliferative diseases, and some developed nothing of note. Apparently the individual “immunity” of the host was an important factor in terms of what response the cancer microbe would elicit.
Tuberculosis infection can affect many parts of the body. Tuberculosis confined to the skin is very different disease when compared to TB of the lung or of the bone. Yet, all three manifestations of the disease are linked together because the TB germ can be found in all three. If cancer microbes are indeed proven as infectious agents in cancer – then various forms of cancer may indeed be manifestations of the same cancer microbe.
There are many “factors” that determine whether a person will become infected with TB. Obviously, smoking is a big factor in lung cancer, radiation is a big factor in skin cancer and leukemia, and so on. However, in defense of the cancer microbe theory, it would be fair to suggest that anything that damages tissue would provide a soil for the possible development of cancer microbe activity in the tissue that could lead to cancer or the development of degenerative or proliferative disease.
Finally, is cancer contagious? For a century physicians have said “no.” But now we know that certain viruses like HIV can lead to cancer. Certain wart “papilloma” viruses can be spread sexually and result in cervical cancer. If further infectious agents, like cancer microbes, are found in cancerous diseases, we may have to reevaluate the contagiosity of cancer.
Obviously in this short communication, few people will be convinced that bacteria cause cancer. For me, it took many years of study, microscopic observation, and communication with microbiologists, pathologists, and colleagues, to become convinced that Livingston and her associates were correct in their claims of a cancer microbe.
A wealth of knowledge pertaining to the cancer microbe (both pro and con) can be found on search engines such as www.google.com. Simply type in “cancer microbe”, “alan cantwell”, “virginia livingston”, “Eleanor Alexander-Jackson” and other names mentioned in this communication.
For a list of scientific publications in medical journals pertaining to the microbiology of cancer, go to the Pubmed website (www.ncbi.nlm.nih.gov) and type in “Cantwell AR”, “Livingston VW”, “Alexander-Jackson E”, “Diller IC”, “Seibert FB.”
For serious students of the microbiology of cancer, I would recommend the following books:
Cantwell, Alan: The Cancer Microbe (1990), Aries Rising Press, Los Angeles
Cantwell, Alan: AIDS: The Mystery and the Solution (1986), Aries Rising Press
Livingston, Virginia: Cancer: A New Breakthrough (1972), Livingston Clinic, San Diego
Livingston, Virginia: The Microbiology of Cancer (1977), Livingston Clinic
Hess, David: Can Bacteria Cause Cancer (1997), NY University Press
Mattman, Lida: Cell Wall Deficient Forms; Stealth Pathogens (1993), CRC Press
Reich, Wilhelm: The Cancer Biopathy (1973), Farrar, Straus, & Giroux, New York
0 comments:
Post a Comment